


Azenta安升达
入驻年限:6 年
- 联系人:
安升达
- 所在地区:
江苏
- 业务范围:
技术服务、细胞库 / 细胞培养、论文服务、试剂、原辅料包材、实验室仪器 / 设备、书籍 / 软件、耗材、医疗器械
- 经营模式:
生产厂商



公司新闻/正文
距离产生美——浅谈寡核苷酸 spacer 修饰的作用
人阅读 发布时间:2023-02-24 11:43
提到寡核苷酸的 spacer 修饰,很多小伙伴可能比较陌生,它们不像荧光、硫代、biotin 修饰等赋予寡核苷酸显而易见的特点;但是在许多生物、化学应用中,使用合适的 spacer,会帮助我们收获更漂亮的结果。本文将给大家分享常见的 spacer 种类与应用实例,让我们共同学习、探讨 spacer 的使用。
什么是 spacer
Spacer 指的是间臂类修饰,大部分 spacer 由直碳链或乙二醇组成。将 spacer 引入到寡核苷酸中,通常是为了在寡核苷酸之间或寡核苷酸与其他官能团之间建立一段距离,以避免空间位阻、减少基团之间不利的相互作用、增加柔性等;或者在不需要寡核苷酸做延伸的情况中,spacer 则作为封闭基团使用。不同 spacer 的原子数量不同,可以通过调节所插入 spacer 的数量与种类达到需要的空间距离。常见的 spacer 有疏水性的 Spacer C3、C6、C12,亲水性的 Spacer 9、Spacer 18,以及特殊的 dSpacer、PC linker。
spacer 种类介绍与应用

Spacer C3 应用实例:
1)Spacer C3 可用于模仿核糖的 3' 和 5' 羟基间的三碳间隔, 因此可用来 「替代」 一段序列中未知的碱基,该应用多见于蛋白与核酸相互作用的研究。
有研究在探索 HIV Tat 蛋白与 RNA 相互作用时,将 RNA 序列中某些核苷酸位点替代为 Spacer C3,从而判断该核苷酸位点在 Tat 蛋白识别 RNA 中是提供结合位点或是作为间隔 [1]。
2)Spacer C3 经常被放在寡核苷酸的 3' 端,作为寡核苷酸延伸的封闭基团,同时 Spacer C3 对于核酸外切酶有一定的抗性。
经典的例子可见于 TaqMan qPCR。TaqMan 探针通常将荧光基团放在探针 5' 端,淬灭基团放在探针 3' 端(更多荧光探针的介绍请点击《修饰基团介绍(荧光探针篇)》),但是在 TaqMan 探针较长时,为了达到较好的淬灭效果,可通过 dT 将淬灭基团引入到探针中间;TaqMan 探针的 3' 端暴露出来,探针可能作为引物在模板上进行非预期的扩增,此时通常会在探针 3' 端修饰 Spacer C3 以封闭探针扩增。类似的,在许多测序引物中也会引入 Spacer C3 起到封闭作用。
图注:F 为荧光基团,Q 为淬灭基团
3)Spacer C3 也可引入到固相载体与寡核苷酸之间,从而减少两者间的空间位阻,有利于寡核苷酸与目标片段的杂交。

Spacer C6 应用实例:
Spacer C6 可引入到固相载体与寡核苷酸之间,从而减少两者间的空间位阻,有利于寡核苷酸与目标片段的杂交。
Spacer C12 应用实例:
Spacer C12 可引入到固相载体与寡核苷酸之间,从而减少两者间的空间位阻,有利于寡核苷酸与目标片段的杂交。
Spacer 9 应用实例:
Spacer 9 常用于桥连两个结构域,赋予两个结构域之间较大的灵活性,有利于结构域发挥作用。比如在 peptide nucleic acid(PNA,肽核酸,一种以多肽骨架取代糖磷酸主链的 DNA 类似物,属于人工产物,主要应用于 DNA 分子的识别与操纵,也有应用于反义核酸药物研究)领域,有研究在 PNA 与 donor DNA 之间使用多个 Spacer 9 做连接。在药物研发中,也有 Spacer 9 连接寡核苷酸与小分子抗原表位的案例 [2]。

Spacer 18 应用实例:
1)Spacer 18 有较好的柔性,常用于辅助核酸链形成发卡结构。在早期已有研究表明,在两段互补寡核苷酸链中引入 Spacer 18 辅助形成的发卡结构,比引入 4 个 T 形成的发卡结构更加稳定 [3]。
2)Spacer 18 作为非核酸的柔性 linker 替代一段核酸序列,提高寡核酸各部分间的灵活性,同时 Spacer 18 不会与体系中其他核酸链产生互补配对。经典的例子就是蝎形探针。
图注:F 为荧光基团,Q 为淬灭基团
3)Spacer 18 常用于引入一个疏水基团,隔开不同的基团以减少不利的位阻或相互作用 [4]。类似的,Spacer 18 也会添加到固相载体与寡核苷酸之间以减少两者间的空间位阻。
例如在 PROER-seq 技术中(通过测序在转录组尺度上揭示蛋白相互作用)[4],作者以 4 个 Spacer 18 为间臂,再利用点击化学技术,在寡核苷酸上添加一个 puromycin(嘌呤霉素)。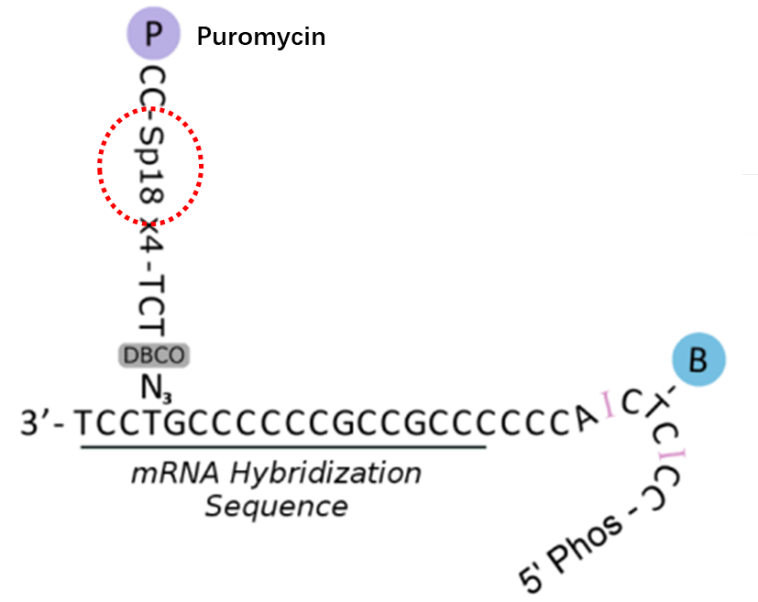
图片来源:《Revealing protein-protein interactions at the transcriptome scale by sequencing》
dSpacer 应用实例:
1)在前文我们提到 Spacer C3 可用于「替代」一个序列中未知的碱基。但是烷基的柔性可能使得糖-磷酸骨架扭曲,使用 dSpacer 则可以缓解这种情况。从 dSpacer 的化学结构我们也可以发现,将 dSpacer 添加到寡核苷酸中,可以引入一个稳定的无碱基位点,模拟体内脱嘌呤导致的基因突变,因此 dSpacer 经常见于 DNA 损伤与修复的研究 [5]。
2)有研究利用 dSpacer 模拟无碱基位点的特性,将 dSpacer 添加到 siRNA 中以降低 siRNA 的脱靶效应。siRNA 是敲低基因的常用手段,但 siRNA 可能作为 miRNA 起作用,从而导致 miRNA-like 的脱靶。研究发现用 dSpacer 取代 siRNA 序列的 pivot 位置(position 6,miRNA 与靶标相互作用的关键位点),可降低 siRNA 的 miRNA-like 脱靶效应,而保留 80%~100% 的 on-target 基因沉默能力 [6]。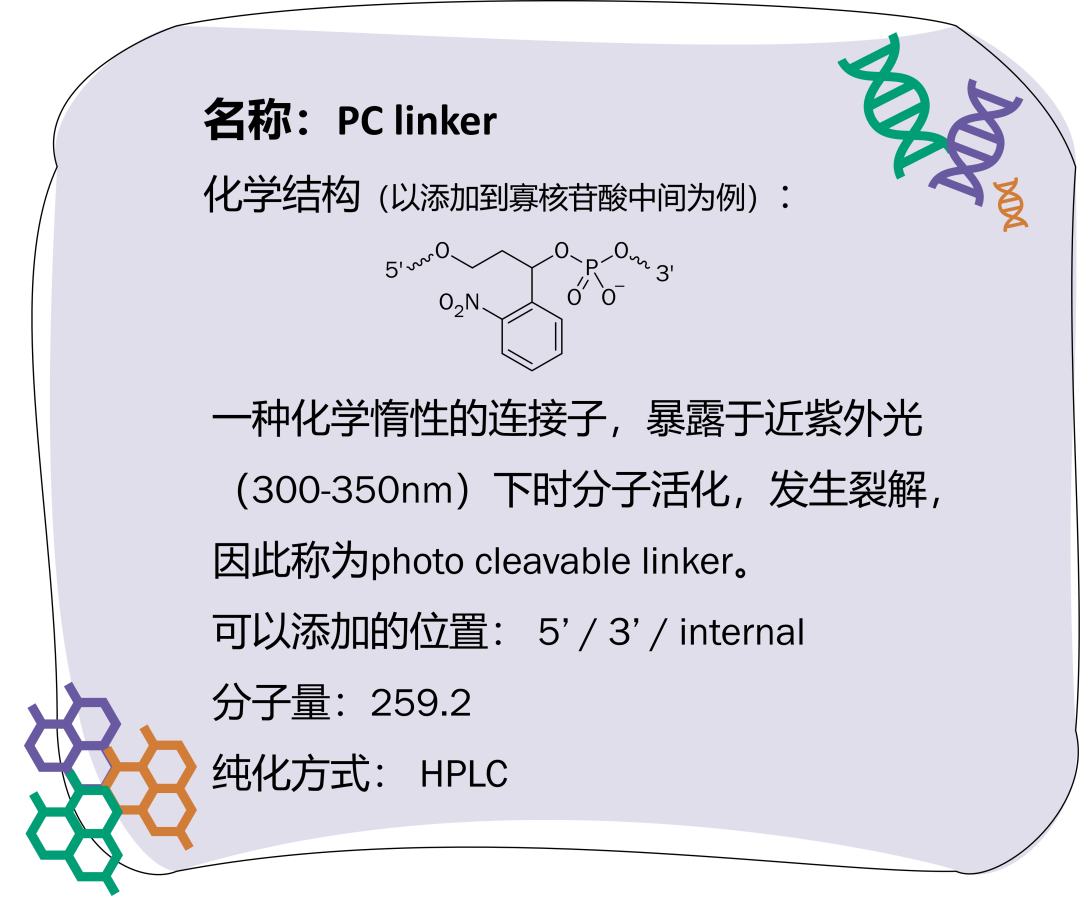
PC linker 应用实例:
PC linker 现广泛应用于分子检测。在 DNA/RNA 的分析中,会给杂交探针添加荧光、生物素、酶等去标记特定的 DNA/RNA 或者作为测序引物。寡核苷酸共价标记的缺点之一是共价键不能被选择性切除,因此标记不能被去除或核酸不能被释放。采用化学方法裂解可能对样品造成影响,且会有化学试剂残留。PC linker 较好地解决了上述难题。
经典的应用可以联想到近几年大热的空间转录组技术。比如 CosMx SMI,该技术用于构建单细胞与亚细胞分辨率的 3D RNA 图谱,通过使用荧光分子 barcode 对生物样本的核酸进行多轮杂交,实现生物样本上亚细胞级分辨率的 RNA 原位检测。如示意图所示,通过在荧光探针上引入 PC site(即 PC linker),新一轮杂交前通过光照射去除上一轮的荧光探针,在不损坏样品的前提下,保证每轮杂交的低背景。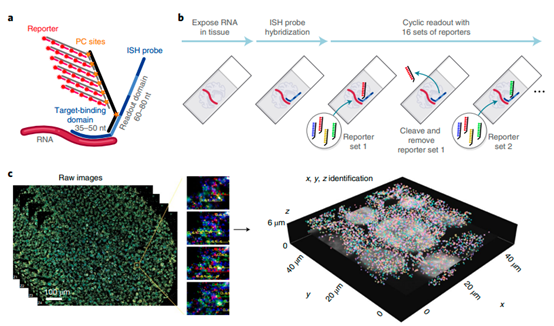
图片来源《High-plex imaging of RNA and proteins at subcellular resolution in fixed tissue by spatial molecular imaging》
看完这些实例,小编感觉寡核苷酸世界丰富多彩,我们可以通过修饰基团赋予寡核苷酸很多功能,但没有 spacer,这些修饰基团也难以发挥最大的作用。当然,spacer 的作用远不止上述的内容,本文仅是抛砖引玉,更多的应用等待大家发掘,欢迎大家一起探讨。
总结表如下:
安升达间臂类修饰引物优势:
以封阻引物(Spacer C3 修饰)为例:
更多寡核苷酸知识分享,欢迎大家持续关注安升达生命科学,安升达引物合成将一直为大家的研究保驾护航!
参考文献:
[1] Pritchard, C. E., Grasby, J. A., Hamy, F., Zacharek, A. M., Singh, M., Karn, J., & Gait, M. J. (1994). Methylphosphonate mapping of phosphate contacts critical for RNA recognition by the human immunodeficiency virus tat and rev proteins. Nucleic acids research, 22(13), 2592–2600. https://doi.org/10.1093/nar/22.13.2592
[2] Palma, E., Klapper, D. G., & Cho, M. J. (2005). Antibodies as drug carriers III: design of oligonucleotides with enhanced binding affinity for immunoglobulin G. Pharmaceutical research, 22(1), 122–127. https://doi.org/10.1007/s11095-004-9017-z
[3] Salunkhe, M., Wu, T., & Letsinger, R. L. (1992). Control of folding and binding of oligonucleotides by use of a nonnucleotide linker. Journal of the American Chemical Society, 114(23), 8768-8772. https://pubs.acs.org/doi/10.1021/ja00049a003
[4] Johnson, K. L., Qi, Z., Yan, Z., Wen, X., Nguyen, T. C., Zaleta-Rivera, K., Chen, C. J., Fan, X., Sriram, K., Wan, X., Chen, Z. B., & Zhong, S. (2021). Revealing protein-protein interactions at the transcriptome scale by sequencing. Molecular cell, 81(19), 4091–4103.e9. https://doi.org/10.1016/j.molcel.2021.07.006
[5] Cheng, K., Xu, Y., Chen, X., Lu, H., He, Y., Wang, L., & Hua, Y. (2020). Participation of RecJ in the base excision repair pathway of Deinococcus radiodurans. Nucleic acids research, 48(17), 9859–9871. https://doi.org/10.1093/nar/gkaa714
[6] Lee, H. S., Seok, H., Lee, D. H., Ham, J., Lee, W., Youm, E. M., Yoo, J. S., Lee, Y. S., Jang, E. S., & Chi, S. W. (2015). Abasic pivot substitution harnesses target specificity of RNA interference. Nature communications, 6, 10154. https://doi.org/10.1038/ncomms10154
[7] He, S., Bhatt, R., Brown, C., Brown, E. A., Buhr, D. L., Chantranuvatana, K., Danaher, P., Dunaway, D., Garrison, R. G., Geiss, G., Gregory, M. T., Hoang, M. L., Khafizov, R., Killingbeck, E. E., Kim, D., Kim, T. K., Kim, Y., Klock, A., Korukonda, M., Kutchma, A., … Beechem, J. M. (2022). High-plex imaging of RNA and proteins at subcellular resolution in fixed tissue by spatial molecular imaging. Nature biotechnology, 10.1038/s41587-022-01483-z. Advance online publication. https://doi.org/10.1038/s41587-022-01483-z
询价列表

暂时没有已询价产品
